Много упреков в адрес селекционеров от любителей поэкспериментировать с пчелами в том, что селекционеры не раскрывают селекционных схем или алгоритмов. Селекция это конечно не игры, но попробуем раскрыть любителям одну из схем – схему двойных межлинейных гибридов.
Итак, для того чтобы селекция и племенной подбор осуществлялись наиболее эффективно, необходимо придерживаться определенной программы разведения. При оценке вашей программы разведения помните, что большинство качеств, которые вы ищете, не могут быть изменены, исправлены или созданы в одном поколении. Чем больше информации вы можете получить о том, как определенные признаки были переданы предками ваших пчел, тем лучше вы можете определить приоритеты и цели разведения. Десятки тысяч генов взаимодействуют, чтобы произвести одну единственную пчелу. Все гены наследуются парами, один ген пары от отца и один от матери. Если пара унаследованных генов от обоих родителей, образно говоря, одинакова, пара называется гомозиготной. Если гены в паре не похожи друг на друга, пара называется гетерозиготной. К счастью, пары генов или вернее набор хромосом, которые делают вид медоносной пчелы медоносной, а не к примеру, восковой или гигантской, всегда гомозиготны и зафиксированы. Аналогично, пары генов, которые делают определенную породу, истинным представителем породы, также гомозиготны и «зафиксированы». Поэтому в каждой породе существует некоторая доля гомозиготных непеременных пар - те, которые дают породе свой специфический стандарт. Тот факт, что селекционер на самом деле не признает существование пород в пчеловодстве, говорит лишь о том, что он понимает, что таких непеременных пар очень мало. Но в так называемых породах много «зафиксированных» пар генов. И все старания селекционеров «чистопородного» разведения заключены в том, чтобы не выйти за рамки таких пар генов и не разрушить эту фиксацию. Ведь такое разрушение чревато негативными последствиями и ведет к усилению генетического груза, которое в последствии очень трудно устранить. Различия между породами выражается в различиях между зафиксированными парами генов этих пород и называется генетическим расстоянием. Пары изменяемых генов, такие как те, которые контролируют фенотип или поведение пчел, производят вариации внутри пород. Такое свойство меняться называется генетической изменчивостью. Большая генетическая изменчивость позволяет передавать нужные качества потомству.
Генетический «мусор», или генетический груз.
Хромосома несет в себе огромное количество генов, которые распределяются на ней линейно, каждый на своем месте. В процессе образования нового организма каждая хромосома материнской и отцовской клеток «отправляет» на слияние свою копию. Так, первая материнская хромосома присоединяется к отцовской хромосоме того же порядка. Гены, расположенные на одном и том же участке хромосом, называются аллельными. Они отвечают за одни и те же наследственные признаки. При этом два одинаковых гена одновременно проявиться не могут, поэтому у конкретного индивида проявляется ген только одной из двух аллелей. Более «сильный» ген подавляет проявление другого – это доминантный ген. Менее «сильный», или образно говоря «сломанный» ген – это рецессивный ген. Проявление генетической изменчивости всегда сопровождается увеличением частоты таких «сломанных» генов, потому что при слиянии отцовских и материнских хромосом более слабые гены «ломаются» и прикрепляются к более сильному гену принимая его «форму» и усиливая его. Хромосомы как бы «притираются» друг к другу, что приводить к разрыву многих аллельных связей, а такие разрывы создают много «сломанных» генов. И чем больше различие или генетическое расстояние в сливающихся хромосомах, тем больше сломанных генов. В первом поколении такие сломанные гены усиливают признаки сильных, но в последующих поколениях они накапливаются и создают генетическое загрязнение или генетическую нагрузку. Но иногда при таких слияниях и нарушениях сцеплений образуются новые уникальные сочетания признаков или кроссенговер. Кроссинговер - это генетическая рекомбинация. Чем равноудалены друг от друга гены, тем чаще между ними происходит кроссинговер. На этом основано проявление выдающихся признаков межпородных гибридов. Но нужно учесть, что генетического загрязнения при кроссенговере образуется ничуть не меньше чем при проявлении любой другой изменчивости, а это значить увеличение генетической нагрузки. Без последующего очищающего отбора (клининга) в этом случае уже не обойтись. Здесь стоить отметить что использование слова "загрязнение" не всегда уместно. Под загрязнением мы понимаем увеличение количества носителей рецессивных генов с неблагоприятными для нас признаками. Но иногда рецессивный признак - это цель селекционера. (В данном контексте мы не имеем ввиду некодирующие функции ДНК в хромосоме или мусорная ДНК - это ДНК, которая не имеет биологически значимой функции, к примеру, псевдогены и фрагменты некогда активных транспозонов).
Теперь, когда мы имеем представление, хотя и сильно упрощенное, о механизмах, протекающих при гибридных скрещиваниях, рассмотрим одну из схем селекции.
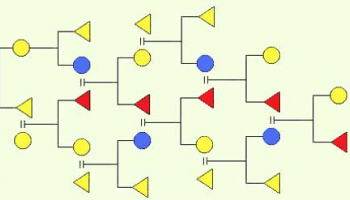
Схема двойных межлинейных гибридов – применяется для слияния ценных селективных признаков двух или четырех инбредных линий.
Двойные гибриды - гибриды, полученные в результате двойной межлинейной гибридизации. Выведение двойных гибридов происходит по следующим схемам:
1. Из четырех инбредных линий А, В, С и Д получают два простых гибрида А х В и С х Д. В результате следующего, двойного межлинейного скрещивания выводят двойных гибридов (АВ)х(СД). Таким образом, двойные гибриды сочетают гены четырех инбредных линий.
2. В том случае если мы с самого начала располагаем только представителями двух инбредных линий А и В , применяется следующая схема. Матку от линии А покрывают трутнем от линии В (А х В). Так же матку от линии В покрывают трутнем от линии А (В х А), и получают два простых межлинейных гибрида АВ и ВА. В результате двойного межлинейного скрещивания выводят двойных гибридов (АВ)х(ВА). Таким образом, двойные гибриды сочетают гены двух инбредных линий.
Двойные гибриды по своим продуктивным качествам превосходят или не уступают межлинейным гибридам первого поколения. При гибридизации по второй схеме мы пользуемся свойствами пространственной ориентации генов, что позволяет нам наложить или объединить признаки с минимальным генетическим мусором. В дальнейшем, от двойных гибридов больше не выводят маток, но используют трутня от них для возвратного скрещивания и приливания новых качеств в исходные инбредные линии.
Выводы: Нужно понимать что эти схемы продуктивны только при использовании исходных инбредных, а следовательно гомозиготных линий. В гетерозиготах же мы получим только накопление генетического мусора, который будет очень трудно вывести. Иногда эти схемы применяются для расширения и закрепления генофонда, ссуженного в ходе созданий инбредных линий. Особенно это касается дизруптивных линий от «однодроновых» маток. Теперь, получив новые знания, мы будем с улыбкой вспоминать, как когда-то без всякого осмысления ломали генетику бакфаста бессмысленными «кроссами» :-).
Основы множественной генетики в селекции медоносных пчел. Ассортативное и диссортативное скрещивание. Инбридинг или аутбридинг?
Автор статьи редактор сайта https://beekingdom.ru